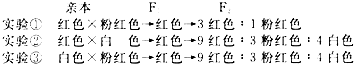19.有两种植物,一种在强光下生长,一种在荫蔽处生长.从这两种植物上各取一片彼此相似的叶片,分别放在两个透明盒子中.在适宜温度条件下,逐渐增加光照强度,测定放氧速率的数据如下表.
(1)由表中数据可以推测,取自强光下的叶片是A;光照强度直接影响光合作用的光反应过程,该过程可将光能转化为ATP、NADPH中的化学能.
(2)光照强度>600μmol光子/(m2•s)时,可推知叶片A放氧速率主要被二氧化碳浓度限制.叶肉细胞呼吸作用产生的CO2转移途径是从线粒体进入叶绿体进行光合作用.
(3)若绘制A、B两叶片放氧速率曲线图,则大约在175μmol光子/(m2•s)时两条曲线相交,此点的生物学含义是阳生植物和阴生植物的净光合速率相等.
(4)为提取叶片B中的色素,研磨前在研钵中除加入剪碎的叶片外,还应加入无水乙醇、碳酸钙、石英砂.经快速研磨、过滤,将滤液收集到试管中,塞上橡皮塞;将试管置于适当的光照条件下2~3min后,试管内的氧含量基本不变.
| 光强[μmol光子/(m2•s)] | 0 | 10 | 25 | 50 | 100 | 250 | 500 | 600 | |
| 放氧速率 [μmolO2/(m2•s)] | 叶片A | -20 | -10 | -5 | -1 | 5 | 15 | 28 | 30 |
| 叶片B | -2 | -0.5 | 1.5 | 3 | 6 | 10 | 12 | 11 | |
(2)光照强度>600μmol光子/(m2•s)时,可推知叶片A放氧速率主要被二氧化碳浓度限制.叶肉细胞呼吸作用产生的CO2转移途径是从线粒体进入叶绿体进行光合作用.
(3)若绘制A、B两叶片放氧速率曲线图,则大约在175μmol光子/(m2•s)时两条曲线相交,此点的生物学含义是阳生植物和阴生植物的净光合速率相等.
(4)为提取叶片B中的色素,研磨前在研钵中除加入剪碎的叶片外,还应加入无水乙醇、碳酸钙、石英砂.经快速研磨、过滤,将滤液收集到试管中,塞上橡皮塞;将试管置于适当的光照条件下2~3min后,试管内的氧含量基本不变.
17.已知番茄植株有茸毛(D)对无茸毛(d)为显性,红果(H)对黄果(h)为显性,两对基因独立遗传.有茸毛番茄植株表面密生茸毛,具有显著的避蚜效果,且能减轻黄瓜花叶病毒的感染,在生产上具有重要的应用价值,但该显性基因纯合时植株不能存活.据此推断错误的是( )
| A. | 将有茸毛番茄白花传粉,子代植株表现型及比例为茸毛:无茸毛=2:1 | |
| B. | 若无茸毛红果番茄与有茸毛黄果番茄杂交,子代出现四种表现型,则子代中不同于亲本表现型的个体占$\frac{1}{2}$ | |
| C. | 若有茸毛红果番茄和有茸毛黄果番茄进行异花传粉,子代只出现了两种表现型,则亲本植株有茸毛红果番茄的基因型仅有DdHH一种 | |
| D. | 若有茸毛红果番茄和有茸毛黄果番茄进行异花传粉,子代只出现了两种表现型,则F1群体中,D的基因频率是$\frac{1}{2}$ |
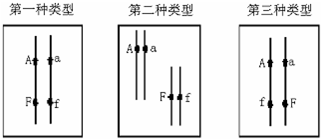
16.如图表示某枝条中生长素相对含量的分布.根据所学知识和图中信息判断,下列叙述错误的是( )
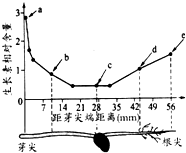

| A. | 茎的向性运动没有体现生长素作用的特点 | |
| B. | a点到c点的方向是根和芽尖极性运输的方向 | |
| C. | b点所对应的细胞物质交换效率比a点所对应的幼苗部位的细胞小 | |
| D. | 如将a点对应浓度的生长素作用于d点细胞,可能会抑制d点细胞的生长 |
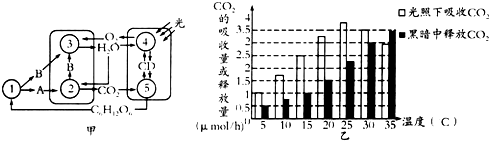
15.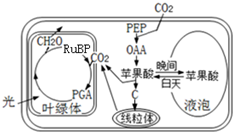 图为菠萝叶肉细胞内的部分代谢示意图,其以气孔白天关闭,晚上开放的特殊方式适应干旱环境.
图为菠萝叶肉细胞内的部分代谢示意图,其以气孔白天关闭,晚上开放的特殊方式适应干旱环境.
(1)图甲所示,PEP、OAA、RuBP、PGA、C为菠萝叶肉细胞内的部分相关代谢物质,能参与CO2固定的有PEP、RuBP(写1个),推测C是丙酮酸.
(2)干旱条件下,菠萝细胞白天产生CO2的具体部位是细胞质基质、线粒体基质;菠萝细胞夜间pH下降,原因是(夜间菠萝细胞合成苹果酸写出1点).
(3)以测定CO2吸收速率与释放速率为指标,探究温度对某绿色植物光合作用与细胞呼吸的影响,结果如下表所示.
①温度在25~30℃间光合作用制造的有机物总量逐渐增加(增加/减少).
②将该植物置于较弱光照下一段时间后取其叶片进行色素分离,与适宜光照下分离的色素带进行比较,发现弱光下滤纸条下端两条色素带明显加宽,推测该植物可通过增加叶绿素含量 以增强对弱光的适应能力.
 图为菠萝叶肉细胞内的部分代谢示意图,其以气孔白天关闭,晚上开放的特殊方式适应干旱环境.
图为菠萝叶肉细胞内的部分代谢示意图,其以气孔白天关闭,晚上开放的特殊方式适应干旱环境.(1)图甲所示,PEP、OAA、RuBP、PGA、C为菠萝叶肉细胞内的部分相关代谢物质,能参与CO2固定的有PEP、RuBP(写1个),推测C是丙酮酸.
(2)干旱条件下,菠萝细胞白天产生CO2的具体部位是细胞质基质、线粒体基质;菠萝细胞夜间pH下降,原因是(夜间菠萝细胞合成苹果酸写出1点).
(3)以测定CO2吸收速率与释放速率为指标,探究温度对某绿色植物光合作用与细胞呼吸的影响,结果如下表所示.
| 温度/℃ | 5 | 10 | 20 | 25 | 30 | 35 |
| 光照条件下CO2吸收速率/(mg•h-1) | 1 | 1.8 | 3.2 | 3.7 | 3.5 | 3 |
| 黑暗条件下CO2释放速率/(mg•h-1) | 0.5 | 0.75 | 1 | 2.3 | 3 | 3.5 |
②将该植物置于较弱光照下一段时间后取其叶片进行色素分离,与适宜光照下分离的色素带进行比较,发现弱光下滤纸条下端两条色素带明显加宽,推测该植物可通过增加叶绿素含量 以增强对弱光的适应能力.
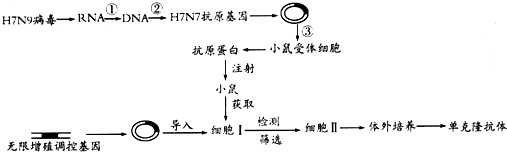
11. 请回答下列与果蝇有关的问题:
请回答下列与果蝇有关的问题:
(1)有一个自然繁殖、表现型正常的果蝇种群,性别比例偏离较大.研究发现该种群的基因库中存在隐性致死突变基因a(胚胎致死).从该种群中选取一对雌雄果蝇相互交配,F1中有202个雌性个体和98个雄性个体.分析可知,F1中雄果蝇的基因型为XAY.若让F1中雌雄果蝇随机交配,F2中雌雄比例是4:3.
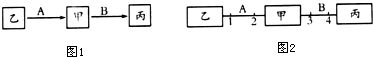
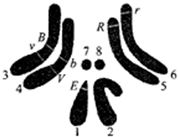
(2)已知四对相对性状中红眼(E)对白眼(e)、灰身(B)对黑身(b)、长翅(V)对残翅(v)、细眼(R)对粗眼(r)为显性.如图是雄果蝇M的四对等位基因在染色体上的分布.
①在雄果蝇M正常减数分裂过程中,含有两条Y染色体的细胞叫做次级精母细胞,欲测定果蝇基因组的序列,需对其中的5条染色体进行DNA测序.
②若果蝇M与基因型为XEXe的个体杂交,子代的雄果蝇眼睛表现型比例为1:1;若果蝇M与黑身残翅个体测交,出现相同比例的灰身长翅和黑身残翅后代,则表明果蝇M在产生配子过程中V和v(或B和b)基因随非姐妹染色单体的交换而发生交换,导致基因重组,产生新的性状组合.
(3)在用基因型为BBvvRRXeY和bbVVrrXEXE的有眼亲本进行杂交获取果蝇M的同时,发现了一只无眼雌果蝇.为分析无眼基因的遗传特点,将该无眼雌果蝇与果蝇M杂交,F1性状分离比如下:
①从实验结果推断,果蝇无眼基因位于7、8(或7、或8)号(填写图中数字)染色体上,理由是无眼、有眼基因与其他各对基因间的遗传均遵循自由组合定律.
②以F1果蝇为材料,设计一步杂交实验判断无眼性状的显隐性.亲本的表现型:F1中的有眼雌雄果蝇.
0 134599 134607 134613 134617 134623 134625 134629 134635 134637 134643 134649 134653 134655 134659 134665 134667 134673 134677 134679 134683 134685 134689 134691 134693 134694 134695 134697 134698 134699 134701 134703 134707 134709 134713 134715 134719 134725 134727 134733 134737 134739 134743 134749 134755 134757 134763 134767 134769 134775 134779 134785 134793 170175
 请回答下列与果蝇有关的问题:
请回答下列与果蝇有关的问题:(1)有一个自然繁殖、表现型正常的果蝇种群,性别比例偏离较大.研究发现该种群的基因库中存在隐性致死突变基因a(胚胎致死).从该种群中选取一对雌雄果蝇相互交配,F1中有202个雌性个体和98个雄性个体.分析可知,F1中雄果蝇的基因型为XAY.若让F1中雌雄果蝇随机交配,F2中雌雄比例是4:3.
(2)已知四对相对性状中红眼(E)对白眼(e)、灰身(B)对黑身(b)、长翅(V)对残翅(v)、细眼(R)对粗眼(r)为显性.如图是雄果蝇M的四对等位基因在染色体上的分布.
①在雄果蝇M正常减数分裂过程中,含有两条Y染色体的细胞叫做次级精母细胞,欲测定果蝇基因组的序列,需对其中的5条染色体进行DNA测序.
②若果蝇M与基因型为XEXe的个体杂交,子代的雄果蝇眼睛表现型比例为1:1;若果蝇M与黑身残翅个体测交,出现相同比例的灰身长翅和黑身残翅后代,则表明果蝇M在产生配子过程中V和v(或B和b)基因随非姐妹染色单体的交换而发生交换,导致基因重组,产生新的性状组合.
(3)在用基因型为BBvvRRXeY和bbVVrrXEXE的有眼亲本进行杂交获取果蝇M的同时,发现了一只无眼雌果蝇.为分析无眼基因的遗传特点,将该无眼雌果蝇与果蝇M杂交,F1性状分离比如下:
| F1 | 雌性﹕雄性 | 灰身﹕黑身 | 长翅﹕残翅 | 细眼﹕粗眼 | 红眼﹕白眼 |
| $\frac{1}{2}$有眼 | 1﹕1 | 3﹕1 | 3﹕1 | 3﹕1 | 3﹕1 |
| $\frac{1}{2}$无眼 | 1﹕1 | 3﹕1 | 3﹕1 | / | / |
②以F1果蝇为材料,设计一步杂交实验判断无眼性状的显隐性.亲本的表现型:F1中的有眼雌雄果蝇.