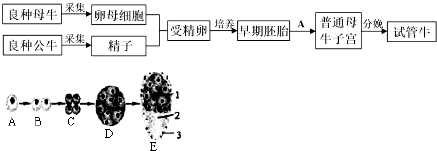
10.初级精母细胞变成次级精母细胞时( )
| A. | 染色体数目减半,DNA 分子数目减半 | |
| B. | 染色体数目减半,DNA 分子数目加倍 | |
| C. | 染色体数目不变,DNA 分子数目加倍 | |
| D. | 染色体数目不变,DNA 分子数目减半 |
9.同源染色体分离、非同源染色体自由组合发生的时期分别是( )
| A. | 减数第一次分裂后期、减数第二次分裂后期 | |
| B. | 减数第一次分裂后期、减数第一次分裂后期 | |
| C. | 减数第二次分裂后期、减数第二次分裂后期 | |
| D. | 有丝分裂后期、有丝分裂后期 |
8.图示为细胞生命历程的概念图.下列有关叙述错误的是( )
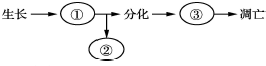
| A. | 细胞表面积与体积的关系限制了细胞的长大 | |
| B. | 过程②中,细胞膜表面的糖蛋白等物质会减少 | |
| C. | 过程③中,细胞的形态、结构和功能均发生改变 | |
| D. | 高度分化的细胞衰老凋亡必将使个体衰老死亡 |
7.油菜物种Ⅰ(2n=20)与Ⅱ(2n=18)杂交产生的幼苗经秋水仙素处理后,得到一个油菜新品系(注:Ⅰ的染色体和Ⅱ的染色体在减数分裂中不会相互配对).
(1)该油菜新品种的培育依据的原理是染色体数目变异.杂交幼苗用秋水仙素处理,导致染色体加倍,获得的植株进行自交,子代不会(会/不会)出现性状分离.
(2)观察油菜新品根尖细胞有丝分裂,处于分裂后期的细胞中含有8个染色体组.
(3)该油菜新品系经过多代种植后出现不同颜色的种子,已知种子颜色由一对基因A/a控制,并受另一对基因R/r影响.用产黑色种子植株(甲)、产黄色种子植株(乙和丙)进行以下实验:
①由实验一得出,种子颜色性状中黑色对黄色为显性.
②分析以上实验可知,两对等位基因之间的相互关系为当R基因存在时会抑制A基因的表达.实验二中F1的基因型为AaRr,F2代产黄色种子植株中能稳定遗传的比例为$\frac{3}{13}$.
③有人重复实验二,发现某一F1植株,其体细胞中含R/r基因的同源染色体有三条(其中两条含r基因),该变异产生的原因分析:由于植株乙在减数第一或二次分裂后期出现了染色体分配异常.让该植株自交(所有配子均有活力),理论上后代中产黑色种子的植株所占比例为$\frac{1}{48}$.
(1)该油菜新品种的培育依据的原理是染色体数目变异.杂交幼苗用秋水仙素处理,导致染色体加倍,获得的植株进行自交,子代不会(会/不会)出现性状分离.
(2)观察油菜新品根尖细胞有丝分裂,处于分裂后期的细胞中含有8个染色体组.
(3)该油菜新品系经过多代种植后出现不同颜色的种子,已知种子颜色由一对基因A/a控制,并受另一对基因R/r影响.用产黑色种子植株(甲)、产黄色种子植株(乙和丙)进行以下实验:
| 组别 | 亲代 | F1表现型 | F1自交所得F2的表现型及比例 |
| 实验一 | 甲×乙 | 全为产黑色种子植株 | 产黑色种子植株:产黄色种子植株=3:1 |
| 实验二 | 乙×丙 | 全为产黄色种子植株 | 产黑色种子植株:产黄色种子植株=3:13 |
②分析以上实验可知,两对等位基因之间的相互关系为当R基因存在时会抑制A基因的表达.实验二中F1的基因型为AaRr,F2代产黄色种子植株中能稳定遗传的比例为$\frac{3}{13}$.
③有人重复实验二,发现某一F1植株,其体细胞中含R/r基因的同源染色体有三条(其中两条含r基因),该变异产生的原因分析:由于植株乙在减数第一或二次分裂后期出现了染色体分配异常.让该植株自交(所有配子均有活力),理论上后代中产黑色种子的植株所占比例为$\frac{1}{48}$.
6.回答下列果蝇眼色的遗传问题.
(1)有人从野生型红眼果蝇中偶然发现一只朱砂眼雄蝇,用该果蝇与一只红眼雌蝇杂交得F1,F1随机交配得F2,子代表现型及比例如下(基因用B、b表示):
①B、b基因位于X染色体上,朱砂眼对红眼为隐性.
②让F2代红眼雌蝇与朱砂眼雄蝇随机交配,所得F3代中,雌蝇有2种基因型,雄蝇中朱砂眼果蝇所占比例为$\frac{1}{4}$.
(2)在实验一F3的后代中,偶然发现一只白眼雌蝇.研究发现,白眼的出现与常染色体上的基因E、e有关.将该白眼果蝇与一只野生型红眼雄蝇杂交得F′1,F′1随机交配得F′2,子代表现型及比例如下:
实验二中亲本白眼雌蝇的基因型为eeXbXb;F′2代杂合雌蝇共有 4种基因型,这些杂合雌蝇中红眼果蝇所占的比例为$\frac{2}{3}$.
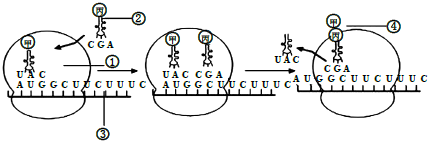
(3)果蝇出现白眼是基因突变导致的,该基因突变前的部分序列(含起始密码信息)如图所示.(注:起始密码子为AUG,终止密码子为UAA、UAG或UGA)

如图所示的基因片段在转录时.若“↑”所指碱基对缺失,该基因控制合成的肽链含5个氨基酸.
(1)有人从野生型红眼果蝇中偶然发现一只朱砂眼雄蝇,用该果蝇与一只红眼雌蝇杂交得F1,F1随机交配得F2,子代表现型及比例如下(基因用B、b表示):
| 实验一 | 亲本 | F1 | F2 | ||
| 雌 | 雄 | 雌 | 雄 | ||
| 红眼(♀)×朱砂眼(♂) | 全红眼 | 全红眼 | 红眼:朱砂眼=1:1 | ||
②让F2代红眼雌蝇与朱砂眼雄蝇随机交配,所得F3代中,雌蝇有2种基因型,雄蝇中朱砂眼果蝇所占比例为$\frac{1}{4}$.
(2)在实验一F3的后代中,偶然发现一只白眼雌蝇.研究发现,白眼的出现与常染色体上的基因E、e有关.将该白眼果蝇与一只野生型红眼雄蝇杂交得F′1,F′1随机交配得F′2,子代表现型及比例如下:
| 实验二 | 亲本 | F′1 | F′2 | |
| 雌 | 雄 | 雌、雄均表现为 红眼:朱砂眼:白眼=4:3:1 | ||
| 白眼(♀)×红眼(♂) | 全红眼 | 全朱砂眼 | ||
(3)果蝇出现白眼是基因突变导致的,该基因突变前的部分序列(含起始密码信息)如图所示.(注:起始密码子为AUG,终止密码子为UAA、UAG或UGA)
如图所示的基因片段在转录时.若“↑”所指碱基对缺失,该基因控制合成的肽链含5个氨基酸.
5.若某研究小组用普通绵羊通过转基因技术获得了转基因绵羊甲和乙各1头,具体见下表.
注:普通绵羊不含A+,B+基因,基因型用A-A-B-B-表示.
请回答:
(1)A+基因转录时,在相关酶的催化下,将游离核苷酸通过磷酸二酯键(化学键名称)聚合成RNA分子. 转基因技术中用到的工具酶有限制酶、DNA连接酶.
(2)为选育黑色细毛的绵羊,分别以绵羊甲、绵羊乙和普通绵羊为亲本杂交获得F1,选择F1中基因型为A+A-B-B-的绵羊和A-A-B+B-的绵羊杂交获得F2.
(3)为获得稳定遗传的黑色细毛绵羊,从F2中选出合适的1对个体杂交得到F3,再从F3中选出2头黑色细毛绵羊(丙、丁)并分析A+和B+基因的表达产物,结果如图所示.不考虑其他基因对A+和B+基因表达产物量的影响,推测绵羊丙的基因型是A+A+B+B-,理论上绵羊丁在F3的黑色细毛绵羊中占的比例是$\frac{1}{9}$.

(4)绵羊甲的体细胞在有丝分裂后期,移向细胞一极的基因组成为A+A-B-B-;绵羊乙的原始生殖细胞在减数第二次分裂后期,移向细胞一极的基因组成为A-B+或A-B-.(注:细胞分裂过程中无基因突变和染色体变异)
0 119775 119783 119789 119793 119799 119801 119805 119811 119813 119819 119825 119829 119831 119835 119841 119843 119849 119853 119855 119859 119861 119865 119867 119869 119870 119871 119873 119874 119875 119877 119879 119883 119885 119889 119891 119895 119901 119903 119909 119913 119915 119919 119925 119931 119933 119939 119943 119945 119951 119955 119961 119969 170175
| 绵羊 | 性别 | 转入的基因 | 基因整合位置 | 表现型 |
| 普通绵羊 | ♀、♂ | - | - | 白色粗毛 |
| 绵羊甲 | ♂ | 1个A+ | 1号常染色体 | 黑色粗毛 |
| 绵羊乙 | ♂ | 1个B+ | 5号常染色体 | 白色细毛 |
请回答:
(1)A+基因转录时,在相关酶的催化下,将游离核苷酸通过磷酸二酯键(化学键名称)聚合成RNA分子. 转基因技术中用到的工具酶有限制酶、DNA连接酶.
(2)为选育黑色细毛的绵羊,分别以绵羊甲、绵羊乙和普通绵羊为亲本杂交获得F1,选择F1中基因型为A+A-B-B-的绵羊和A-A-B+B-的绵羊杂交获得F2.
(3)为获得稳定遗传的黑色细毛绵羊,从F2中选出合适的1对个体杂交得到F3,再从F3中选出2头黑色细毛绵羊(丙、丁)并分析A+和B+基因的表达产物,结果如图所示.不考虑其他基因对A+和B+基因表达产物量的影响,推测绵羊丙的基因型是A+A+B+B-,理论上绵羊丁在F3的黑色细毛绵羊中占的比例是$\frac{1}{9}$.
(4)绵羊甲的体细胞在有丝分裂后期,移向细胞一极的基因组成为A+A-B-B-;绵羊乙的原始生殖细胞在减数第二次分裂后期,移向细胞一极的基因组成为A-B+或A-B-.(注:细胞分裂过程中无基因突变和染色体变异)