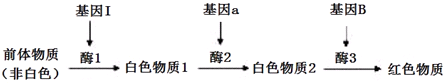
18.某种花卉的野生型 全部开红花,但实验室通过育种得到了两个开白花的突变品系.为了研究该花卉的 花色遗传方式,现用野生型和两个纯种突变品系分别进行杂交
实验并均得到F1,F1自交得F2,结果见表格:
(1)甲同学只观察杂交组合I就推断该花卉的花色由一对等位基因控制.若该假设成立,将所有红花选出进行自交得到的F3中白花基因频率为$\frac{1}{3}$.
(2)乙同学发现杂交组合II的实验结果和甲同学的假设矛盾,于是通过查阅资料发现该花卉的花色由两对位于非同源染色体上的等位基因决定(产生红色素的基因A对a为显性;B对b为显性,其中一个基因的表达能抑制花瓣中所有色素的合成).据此回答下列问题:
①能够抑制花瓣色素产生的基因是B (填“B”或“b”),野生型红花的基因型为AAbb
②杂交组合Ⅲ的F2表现型及比例为红花:白花=3:13.
(3)在(2)成立的前提下,科学家从蓝色三叶草中获取了蓝色素基因M.
①能够将M基因送入该花卉体细胞的工具叫运载体
②为了培育出开纯蓝花的花卉,最好选择基因型为aabb 的花卉体细胞作受体细胞.
③将一个M基因成功整合到细胞的某条染色体上,并通过组织培养得到开蓝色花的植株,为了尽快获得能够稳定遗传的开蓝花品系,可选用的育种方法为单倍体育种.
| 组别 | 亲本 | F1表现 | F2表现 |
| Ⅰ | 突变品系1×野生型 | 全开红花 | $\frac{3}{4}$红花,$\frac{1}{4}$白花 |
| Ⅱ | 突变品系2×野生型 | 全开白花 | $\frac{1}{4}$红花,$\frac{3}{4}$白花 |
| Ⅲ | 突变品系1×突变品系2 | 全开白花 | ? |
(1)甲同学只观察杂交组合I就推断该花卉的花色由一对等位基因控制.若该假设成立,将所有红花选出进行自交得到的F3中白花基因频率为$\frac{1}{3}$.
(2)乙同学发现杂交组合II的实验结果和甲同学的假设矛盾,于是通过查阅资料发现该花卉的花色由两对位于非同源染色体上的等位基因决定(产生红色素的基因A对a为显性;B对b为显性,其中一个基因的表达能抑制花瓣中所有色素的合成).据此回答下列问题:
①能够抑制花瓣色素产生的基因是B (填“B”或“b”),野生型红花的基因型为AAbb
②杂交组合Ⅲ的F2表现型及比例为红花:白花=3:13.
(3)在(2)成立的前提下,科学家从蓝色三叶草中获取了蓝色素基因M.
①能够将M基因送入该花卉体细胞的工具叫运载体
②为了培育出开纯蓝花的花卉,最好选择基因型为aabb 的花卉体细胞作受体细胞.
③将一个M基因成功整合到细胞的某条染色体上,并通过组织培养得到开蓝色花的植株,为了尽快获得能够稳定遗传的开蓝花品系,可选用的育种方法为单倍体育种.
16.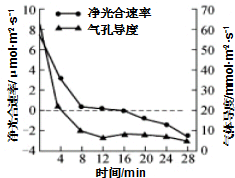 科研人员用茶树离体叶片测定了气孔导度(指单位时间、单位面积叶片通过气孔的气体量)、净光合速率的变化,结果如图所示.下列说法错误的是( )
科研人员用茶树离体叶片测定了气孔导度(指单位时间、单位面积叶片通过气孔的气体量)、净光合速率的变化,结果如图所示.下列说法错误的是( )
 科研人员用茶树离体叶片测定了气孔导度(指单位时间、单位面积叶片通过气孔的气体量)、净光合速率的变化,结果如图所示.下列说法错误的是( )
科研人员用茶树离体叶片测定了气孔导度(指单位时间、单位面积叶片通过气孔的气体量)、净光合速率的变化,结果如图所示.下列说法错误的是( )| A. | 实验开始4min时,水光解产生气体的去向有进入线粒体和释放到细胞外 | |
| B. | 若茶树植株长期处于16min时的状态,则其不能正常生长 | |
| C. | 与8min时相比,4min时叶绿体中的ADP、NADP+含量较少 | |
| D. | 在最初0~4min内,叶肉细胞净光合速率下降的主要原因是气孔导度的下降,阻止了CO2的供应 |
15.研究发现,细胞中染色体的正确排列与分离依赖于染色单体之间的粘连.动物细胞内存在有一种SGO蛋白,保护粘连蛋白不被水解酶破坏,其主要集中在染色体的着丝粒位置,在染色体的其他位置也有分布,如图所示.下列说法错误的是( )
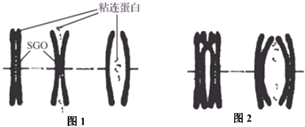

| A. | 图2中的粘连蛋白可能与同源染色体的非姐妹染色单体发生片段交换有关 | |
| B. | 在细胞分裂过程中,细胞会产生水解酶将粘连蛋白分解,而染色体上的其他蛋白质不受影响,这体现了酶的专一性 | |
| C. | 阻断正在分裂的动物体细胞内SGO蛋白的合成,细胞最可能出现染色体数目变异 | |
| D. | 图1所发生的变化只发生在有丝分裂,图2发生的变化只发生在减数分裂 |
14.褐飞虱能吸食水稻汁液,是稻田常见害虫,其成虫具有长翅和短翅两种翅型,短翅雌虫产卵量大、常使虫口激增,长翅型飞行能力强、有利于迁飞.为了解褐飞虱翅型的遗传规律,研究人员进行了以下杂交实验:
(1)实验一和实验二的杂交结果相同(相同/不同),说明控制翅型的基因位于常染色体上.
(2)研究人员认为,褐飞虱翅型受一对等位基因控制(B控制短翅、B′控制长翅).据此分析,实验一和二的子一代雄虫全为长翅、雌虫全为短翅的原因是:子一代基因型都为BB’,雌虫的短翅基因B为显性,雄虫的长翅基因B’为显性,实验三和四的结果支持此假设.①至④个体中,①的基因型与其他不同.
(3)研究人员从稻田中随机捕获若干长翅雌雄褐飞虱自由交配,子一代出现少量短翅雌虫,请根据亲本基因型解释原因:长翅雌虫的基因型为B’B’,长翅雄虫的基因型为BB’或B’B’,子代较小比例的BB’中,只有雌虫表现为短翅.子一代中短翅雌虫的比例取决于所捕获长翅褐飞虱中杂合长翅雄虫的比例(或:B基因频率).若每一代的雌雄个体均自由交配,从子二代开始会出现短翅雄虫.
(4)田间监测表明,褐飞虱长短翅型的数量变化与水稻发育期密切相关,水稻孕穗至开花期有利于短翅型发生,黄熟期有利于长翅型发生,这说明褐飞虱翅型是基因型和环境共同作用的结果.褐飞虱长短翅型的数量变化有利于增强褐飞虱对环境的适应.
| 组别 | 亲本 | 子一代 |
| 实验一 | 纯合短翅雄虫①╳纯合长翅雌虫 | 长翅雄②:短翅雌③=1:1 |
| 实验二 | 纯合长翅雄虫╳纯合短翅雌虫 | 长翅雄:短翅雌④=1:1 |
| 实验三 | 纯合短翅雄虫①╳短翅雌③ | 长翅雄:短翅雄:短翅雌=1:1:2 |
| 实验四 | 长翅雄虫②╳纯合长翅雌虫 | 长翅雄:长翅雌:短翅雌=1:2:1 |
(2)研究人员认为,褐飞虱翅型受一对等位基因控制(B控制短翅、B′控制长翅).据此分析,实验一和二的子一代雄虫全为长翅、雌虫全为短翅的原因是:子一代基因型都为BB’,雌虫的短翅基因B为显性,雄虫的长翅基因B’为显性,实验三和四的结果支持此假设.①至④个体中,①的基因型与其他不同.
(3)研究人员从稻田中随机捕获若干长翅雌雄褐飞虱自由交配,子一代出现少量短翅雌虫,请根据亲本基因型解释原因:长翅雌虫的基因型为B’B’,长翅雄虫的基因型为BB’或B’B’,子代较小比例的BB’中,只有雌虫表现为短翅.子一代中短翅雌虫的比例取决于所捕获长翅褐飞虱中杂合长翅雄虫的比例(或:B基因频率).若每一代的雌雄个体均自由交配,从子二代开始会出现短翅雄虫.
(4)田间监测表明,褐飞虱长短翅型的数量变化与水稻发育期密切相关,水稻孕穗至开花期有利于短翅型发生,黄熟期有利于长翅型发生,这说明褐飞虱翅型是基因型和环境共同作用的结果.褐飞虱长短翅型的数量变化有利于增强褐飞虱对环境的适应.
13.鼠源单克隆抗体会使人产生免疫反应,导致抗体失效,科研人员通过转基因技术对抗体结构进行人源化改造.关于该过程的叙述不正确的是( )
| A. | 将含有改造后抗体基因的重组载体导入鼠受精卵 | |
| B. | 将鼠骨髓瘤细胞与转基因鼠B淋巴细胞融合并筛选杂交瘤细胞 | |
| C. | 用核酸分子杂交技术筛选出能产生目的抗体的杂交瘤细胞 | |
| D. | 在体外对所选杂交瘤细胞进行培养以获取大量人源单克隆抗体 |
12.新疆草原蝗灾主要由3种蝗虫引起,总密度达到20头/m2以上,研究人员利用从尼日尔蝗虫体内分离得到的绿僵菌来做虫害治理实验.喷洒1h后,随机采集蝗虫饲养,每天记录死亡个体数,第7天的实验结果如表:
以下分析正确的是( )
0 118641 118649 118655 118659 118665 118667 118671 118677 118679 118685 118691 118695 118697 118701 118707 118709 118715 118719 118721 118725 118727 118731 118733 118735 118736 118737 118739 118740 118741 118743 118745 118749 118751 118755 118757 118761 118767 118769 118775 118779 118781 118785 118791 118797 118799 118805 118809 118811 118817 118821 118827 118835 170175
| 欧亚草地蝗 | 红胫戟纹蝗 | 西伯利亚蝗 | |
| 第7d死亡率 | 98.8% | 92.2% | 93.6% |
| A. | 本实验采用化学药剂处理蝗虫 | |
| B. | 用绿僵菌治理虫害会引起生物放大 | |
| C. | 绿僵菌专性寄生于尼日尔蝗虫 | |
| D. | 绿僵菌处理会改变蝗虫种群的基因频率 |