6.甘蓝型油菜花色性状由三对等位基因控制,分别位于三对同源染色体上.花色表现型与基因型之间的对应关系如下表.
(1)白花植株的基因型有9种,其中自交不会出现性状分离的基因型有9种.乳白花植株自交能否出现后代全部为乳白花的植株?不能(填“能”或“不能”).
(2)黄花植株同金黄花植株杂交得F1,F1自交后代出现两种表现型且比例为3:1,则黄花植株基因型为aaBBdd或aabbDD.黄花植株自交后代出现金黄花植株的概率最高为$\frac{1}{4}$.
(3)乳白色植株在产生配子时,基因A和a的分离(不考虑交叉互换)发生在时期.基因A和A的分离发生在减数第二次分裂后期时期.
(4)若让基因型为AaBbDd的植株自交,后代将出现4种不同花色的植株,若让其进行测交,则其后代的表现型及其比例为乳白花:黄花:金黄花=4:3:1.
| 表现型 | 白花 | 乳白花 | 黄花 | 金黄花 |
| 基因型 | AA____ | Aa____ | aaB___、aa__D_、aaB_D_ | aabbdd |
(2)黄花植株同金黄花植株杂交得F1,F1自交后代出现两种表现型且比例为3:1,则黄花植株基因型为aaBBdd或aabbDD.黄花植株自交后代出现金黄花植株的概率最高为$\frac{1}{4}$.
(3)乳白色植株在产生配子时,基因A和a的分离(不考虑交叉互换)发生在时期.基因A和A的分离发生在减数第二次分裂后期时期.
(4)若让基因型为AaBbDd的植株自交,后代将出现4种不同花色的植株,若让其进行测交,则其后代的表现型及其比例为乳白花:黄花:金黄花=4:3:1.
5.依靠生态系统的自身调节能力,辅以有效的人为手段,将已经被破坏的生态系统,恢复为健康状态的过程称为生态恢复.该过程中( )
| A. | 发生了初生演替 | B. | 能量的输入大于散失 | ||
| C. | 抵抗力稳定性保持不变 | D. | 恢复为原来的物种组成 |
4.下列与细胞工程技术相关的叙述中,不正确的是( )
| A. | 植物组织培养技术的理论基础是细胞的全能性 | |
| B. | 植物体细胞杂交技术可以克服远缘杂交不亲和障碍 | |
| C. | 动物细胞融合与植物原生质体融合的基本原理相同 | |
| D. | 动物细胞培养与植物组织培养所用的培养基成分基本相同 |
 如图为DNA分子结构模式图,请据图回答:
如图为DNA分子结构模式图,请据图回答:
2. 特异性免疫包括体液免疫和细胞免疫.如图为人的淋巴细胞免疫图解,请据图分析下列说法中正确的是( )
特异性免疫包括体液免疫和细胞免疫.如图为人的淋巴细胞免疫图解,请据图分析下列说法中正确的是( )
 特异性免疫包括体液免疫和细胞免疫.如图为人的淋巴细胞免疫图解,请据图分析下列说法中正确的是( )
特异性免疫包括体液免疫和细胞免疫.如图为人的淋巴细胞免疫图解,请据图分析下列说法中正确的是( )| A. | 细胞②为B细胞 | B. | 细胞⑤产生的物质为球蛋白 | ||
| C. | 细胞⑤为靶细胞 | D. | 细胞③为效应T细胞 |
1.下列有关细胞生命历程的说法不正确的是( )
| A. | 早期胚胎细胞通过细胞分裂分化形成各种组织和器官 | |
| B. | 细胞分化导致基因选择性表达,细胞种类增多 | |
| C. | 癌变的细胞,其膜表面的糖蛋白减少 | |
| D. | 细胞凋亡的过程是受基因控制的 |
19.出芽酵母的生活史如图1所示,其野生型基因发生突变后,表现为突变型(如图2所示).研究发现该突变型酵母(单倍体)中有少量又回复为野生型表现型,请分析回答:
部分密码子表
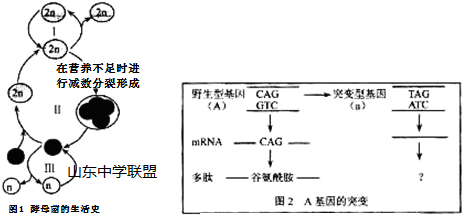
(1)酵母的生殖方式Ⅱ与Ⅰ、Ⅲ相比,在减数分裂过程中能发生基因重组,因而产生的后代具有更大的变异性.
(2)依据图2和表1分析,A基因的突变会导致相应蛋白质的合成提前终止,进而使其功能缺失.
(3)研究者提出两种假设来解释突变型酵母回复为野生型表现型的原因.
①假设一:a基因又突变回A基因.提出此假设的依据是基因突变具有可逆性.
②假设二:a基因未发生突变,编码能携带谷氨酰胺的tRNA的基因B突变为b基因(a、b基因位于非同源染色体上).在a基因表达过程中,b基因的表达产物携带的氨基酸为谷氨酰胺,识别的密码子为UAG,使a基因指导合成出完整的、有功能的蛋白质.
(4)为检验以上假设是否成立,研究者将回复后的单倍体野生型酵母与原始单倍体野生型酵母进行两细胞融合,获取二倍体个体(F1),培养F1,使其减数分裂产生大量单倍体后代,检测并统计这些单倍体的表现型.
①若F1的单倍体子代表现型为全部为野生型,则支持假设一.
②若F1的单倍体子代野生型与突变型比例为3:1,则支持假设二,F1的单倍体子代中野生型个体的基因型是AB、Ab、ab,来源于一个F1细胞的四个单倍体子代酵母细胞的表现型及比例可能为全部为野生型、野生型:突变型=3:1、野生型:突变型=1:1.
0 116028 116036 116042 116046 116052 116054 116058 116064 116066 116072 116078 116082 116084 116088 116094 116096 116102 116106 116108 116112 116114 116118 116120 116122 116123 116124 116126 116127 116128 116130 116132 116136 116138 116142 116144 116148 116154 116156 116162 116166 116168 116172 116178 116184 116186 116192 116196 116198 116204 116208 116214 116222 170175
部分密码子表
| 第一个字母 | 第二个字母 | 第三个字母 | |
| A | U | ||
| U | 终止 | 亮氨酸 | G |
| C | 谷氨酰胺 | 亮氨酸 | G |
| A | 天冬酰胺 | 异亮氨酸 | C |
| G | 谷氨酸 | 氨酸 | A |

(1)酵母的生殖方式Ⅱ与Ⅰ、Ⅲ相比,在减数分裂过程中能发生基因重组,因而产生的后代具有更大的变异性.
(2)依据图2和表1分析,A基因的突变会导致相应蛋白质的合成提前终止,进而使其功能缺失.
(3)研究者提出两种假设来解释突变型酵母回复为野生型表现型的原因.
①假设一:a基因又突变回A基因.提出此假设的依据是基因突变具有可逆性.
②假设二:a基因未发生突变,编码能携带谷氨酰胺的tRNA的基因B突变为b基因(a、b基因位于非同源染色体上).在a基因表达过程中,b基因的表达产物携带的氨基酸为谷氨酰胺,识别的密码子为UAG,使a基因指导合成出完整的、有功能的蛋白质.
(4)为检验以上假设是否成立,研究者将回复后的单倍体野生型酵母与原始单倍体野生型酵母进行两细胞融合,获取二倍体个体(F1),培养F1,使其减数分裂产生大量单倍体后代,检测并统计这些单倍体的表现型.
①若F1的单倍体子代表现型为全部为野生型,则支持假设一.
②若F1的单倍体子代野生型与突变型比例为3:1,则支持假设二,F1的单倍体子代中野生型个体的基因型是AB、Ab、ab,来源于一个F1细胞的四个单倍体子代酵母细胞的表现型及比例可能为全部为野生型、野生型:突变型=3:1、野生型:突变型=1:1.
