题目内容
9.染色体为AaBb的个体,正常情况下不可能产生的配子是( )| A. | Aa | B. | AB | C. | aB | D. | ab |
分析 1、基因分离定律的实质:在杂合子的细胞中,位于一对同源染色体上的等位基因,具有一定的独立性;生物体在进行减数分裂形成配子时,等位基因会随着同源染色体的分开而分离,分别进入到两个配子中,独立地随配子遗传给后代.
2、基因自由组合定律的实质是:位于非同源染色体上的非等位基因的分离或自由组合是互不干扰的;在减数分裂过程中,同源染色体上的等位基因彼此分离的同时,非同源染色体上的非等位基因自由组合.
解答 解:A和a、B和b是两对等位基因,在减数第一次分裂后期,等位基因随着同源染色体的分开而分离,同时非同源染色体上的非等位基因之间自由组合,因此,基因型为AaBb的个体正常情况下能产生四种配子,即AB、Ab、ab、aB.
故选:A.
点评 本题考查基因的自由组合定律和细胞的减数分裂,要求考生识记细胞的减数分裂过程,尤其是减数第一次分裂后期的特点;掌握基因的分离定律和基因的自由组合定律,能判断基因型为AaBb的个体产生配子的种类,再选出正确答案即可.

练习册系列答案
相关题目
2.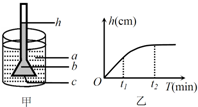 某兴趣小组用图甲研究渗透作用,a和b均为蔗糖溶液,c允许单糖通过.图乙为实验结果.下列说法正确的是( )
某兴趣小组用图甲研究渗透作用,a和b均为蔗糖溶液,c允许单糖通过.图乙为实验结果.下列说法正确的是( )
某兴趣小组用图甲研究渗透作用,a和b均为蔗糖溶液,c允许单糖通过.图乙为实验结果.下列说法正确的是( )| A. | 烧杯中蔗糖溶液浓度大于漏斗中蔗糖溶液浓度 | |
| B. | t2时刻半透膜两侧蔗糖溶液浓度相同 | |
| C. | 若t2时刻漏斗中加入蔗糖酶,则h将先上升后下降 | |
| D. | 若t2时刻烧杯中加入蔗糖酶,则h最终将下降 |
19.蓝藻与酵母菌的相同之处是( )
| A. | 都有拟核 | B. | 都有线粒体 | ||
| C. | 都能进行光合作用 | D. | 都能进行有氧呼吸 |
5..
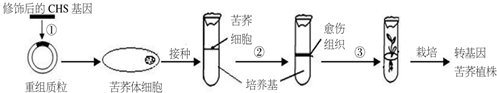
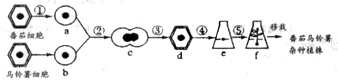
(1)图中常用的是MS培养基.实验室一般使用保存的培养基母液来制备该培养基,配置母液时无机物中的大量元素浓缩10倍,激素、维生素等有机物单独制成1mg/mL的母液,接种前,用灭菌后未接种的培养基培养一段时间,观察是否形成菌落,目的是判断培养基灭菌是否彻底.
(2)过程②、③分别是脱分化和再分化.苦荞体细胞能培养成植株说明植物细胞具有全能性.
(3)为探究激素对于过程诱导愈伤组织的影响,某研究小组在培养基中加入6-BA和2,4─D,灭菌后分别接种,在适宜条件下培养一段时间后,统计愈伤组织的诱导率,实验结果如下表:
该实验的因变量是诱导率,诱导的最佳组合是0.1mg•L-16-BA和4mg•L-12,4-D.
(4)判断转基因苦荞培育是否成功,可比较转基因苦荞与普通苦荞的黄酮类化合物含量,也可测定细胞中CHS含量.
(1)图中常用的是MS培养基.实验室一般使用保存的培养基母液来制备该培养基,配置母液时无机物中的大量元素浓缩10倍,激素、维生素等有机物单独制成1mg/mL的母液,接种前,用灭菌后未接种的培养基培养一段时间,观察是否形成菌落,目的是判断培养基灭菌是否彻底.
(2)过程②、③分别是脱分化和再分化.苦荞体细胞能培养成植株说明植物细胞具有全能性.
(3)为探究激素对于过程诱导愈伤组织的影响,某研究小组在培养基中加入6-BA和2,4─D,灭菌后分别接种,在适宜条件下培养一段时间后,统计愈伤组织的诱导率,实验结果如下表:
| 实验编号 | 浓度/mg•L-1 | 诱导率% | 实验编号 | 浓度/mg•L-1 | 诱导率% | 实验编号 | 浓度/mg•L-1 | 诱导率% | |||
| 2,4-D | 6-BA | 2,4-D | 6-BA | 2,4-D | 6-BA | ||||||
| 1 | 0 | 0 | 0 | 5 | 0 | 0.1 | 6 | 9 | 0 | 0.2 | 5 |
| 2 | 2 | 35 | 6 | 2 | 91 | 10 | 2 | 60 | |||
| 3 | 4 | 65 | 7 | 4 | 100 | 11 | 4 | 80 | |||
| 4 | 6 | 40 | 8 | 6 | 93 | 12 | 6 | 72 | |||
(4)判断转基因苦荞培育是否成功,可比较转基因苦荞与普通苦荞的黄酮类化合物含量,也可测定细胞中CHS含量.
14.下列有关细胞生命历程的叙述,不正确的是( )
| A. | 细胞生长使物质交换效率减慢 | |
| B. | 细胞分化、衰老和癌变的共同特征是细胞代谢速率减慢 | |
| C. | 细胞分化是不同细胞遗传信息的执行情况不同导致的 | |
| D. | 细胞全能性的表现必须要通过细胞分化才能实现 |
1.动物细胞工程常用的技术手段中,基础技术是( )
| A. | 动物细胞核移植 | B. | 动物细胞融合 | C. | 单克隆抗体 | D. | 动物细胞培养 |
18.下列属于可表示纯合体的是( )
| A. | BbDd | B. | bbDd | C. | BBXDXD | D. | BbXdY |