题目内容
某种野生型油菜存在一种突变体,叶绿素、类胡萝卜素含量均低,其叶片呈现黄化色泽。野生型和突变体的成熟叶片净光合速率、呼吸速率及相关指标见下表。请分析并回答:
指标 类型 | 类胡萝卜素/叶绿素 | 叶绿素a/b | 净光合速率 (μmolCO2•m-2•s-1) | 胞间CO2浓度 (μmolCO2•m-2•s-1) | 呼吸速率 (μmolCO2•m-2•s-1) |
野生型 | 0.28 | 6.94 | 8.13 | 210.86 | 4.07 |
突变体 | 0.32 | 9.30 | 5.66 | 239.07 | 3.60 |
(1)类胡萝卜素主要吸收 光;欲测定色素含量,可选用 (溶剂)提取叶片中的色素。
(2)据测定,突变体处于发育初期的叶片,类胡萝卜素与叶绿素的比值为0.48,可以推测,叶绿素在色素组成中所占的比例随叶片发育逐渐 。与野生型相比,突变体发生的改变可能 (促进/抑制)叶绿素a向叶绿素b的转化。
(3)突变体成熟叶片中叶绿体消耗CO2的速率比野生型低 μmolCO2•m-2•s-1。经分析,研究人员认为CO2浓度不是导致突变体光合速率降低的限制因素,依据是 。


 名校联盟快乐课堂系列答案
名校联盟快乐课堂系列答案(1)秋水仙素通过抑制分裂细胞中纺锤体的形成,导致染色体加倍.
(2)观察油菜新品系根尖细胞有丝分裂,处于分裂后期的细胞中含有76条染色体.
(3)该油菜新品系经多代种植后出现不同颜色的种子,已知种子颜色由一对基因A/a控制,并受另一对基因R/r影响.用产黑色种子植株(甲)、产黄色种子植株(乙和丙)进行以下实验:
| 组别 | 亲代 | F1表现型 | F1自交所得F2的表现型及比例 |
| 实验一 | 甲×乙 | 全为产黑色种子植株 | 产黑色种子植株:产黄色种子植株=3:1 |
| 实验二 | 乙×丙 | 全为产黄色种子植株 | 产黑色种子植株:产黄色种子植株=3:13 |
②分析以上试验可知,当R基因存在时会抑制A基因的表达.实验二中丙的基因型为AARR,F2代产黄色种子植株中杂合子的比例为$\frac{10}{13}$.
③有人重复实验二,发现一F1植株,其体细胞中含R/r基因的同源染色体有三条(其中两条含R基因),请解释该变异产生的原因:植株丙在减数第一次分裂后期含R基因的同源染色体未分离.让该植株自交,理论上后代中产黑色种子的植株所占比例为$\frac{1}{48}$.
下表为某学校兴趣小组探究影响植物光合作用因素所做的实验。
容器 | 植物 | 部位 | 光质 | 光照强度 (klx) | 温度(℃) | O2增加速率(mL/8小时) |
1 | 天缝葵 | 叶 | 红 | 20 | 22 | 120 |
2 | 天世葵 | 叶 | 黄 | 20 | 22 | 15 |
3 | 天世葵 | 根 | 红 | 20 | 22 | -10 |
4 | 紫罗兰 | 叶 | 红 | 20 | 22 | 80 |
5 | 紫罗兰 | 叶 | 黄 | 20 | 22 | 10 |
(1)1号容器天竺葵产氧速率是 mL/8小时。
(2)据表可知,探究影响植物光合作用因素有 。
(3)1号容器比2号容器O2增加速率大,只是因为天竺葵吸收红光比黄光多,而不是不同光质下天竺葵呼吸速率不同,为了充分证明这一结论。还要增加6号实验组,这组实验如何设计: 。 若6号容器天竺葵的O2增加速率与 号的相同,则上述结论成立。
(4)5号容器紫罗兰的线粒体内将丙酮酸的能量转化成 。
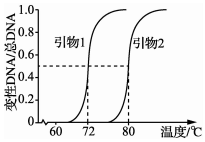 根据某基因上游和下游的碱基序列,设计合成了用于该基因PCR的两段引物(单链 DNA).引物与该基因变性DNA(单链DNA)结合为双链DNA的过程称为复性.如图是两引物的Tm(引物熔解温度,即50%的引物与其互补序列形成双链DNA分子时的温度)测定结果,下列叙述错误的是( )
根据某基因上游和下游的碱基序列,设计合成了用于该基因PCR的两段引物(单链 DNA).引物与该基因变性DNA(单链DNA)结合为双链DNA的过程称为复性.如图是两引物的Tm(引物熔解温度,即50%的引物与其互补序列形成双链DNA分子时的温度)测定结果,下列叙述错误的是( )| A. | 通常引物1和引物2不能有碱基互补配对关系 | |
| B. | 两引物分别是子链延伸的起点,并可以反复利用 | |
| C. | 若两引物的脱氧核苷酸数相同,推测引物2的GC含量较高 | |
| D. | 复性所需温度与时间,取决于引物的长度、碱基组成及其浓度等因素 |
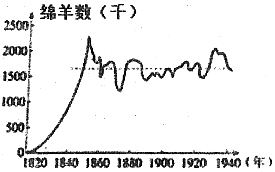
| A. | 1840年前种群数量的增长趋势与气候适宜、食物和空间较充裕有关 | |
| B. | 该绵羊种群的环境容纳量会因环境的破坏而改变 | |
| C. | 种群数量达到环境容纳量后,出生率和死亡率基本相等 | |
| D. | 该绵羊种群的环境容纳量在1850-1940年间不断地波动 |